 细胞代谢及代谢副产物对细胞培养的影响doc.docx
细胞代谢及代谢副产物对细胞培养的影响doc.docx
- 文档编号:15940965
- 上传时间:2023-07-09
- 格式:DOCX
- 页数:13
- 大小:199.71KB
细胞代谢及代谢副产物对细胞培养的影响doc.docx
《细胞代谢及代谢副产物对细胞培养的影响doc.docx》由会员分享,可在线阅读,更多相关《细胞代谢及代谢副产物对细胞培养的影响doc.docx(13页珍藏版)》请在冰点文库上搜索。
细胞代谢及代谢副产物对细胞培养的影响doc
细胞代谢及代谢副产物对细胞培养的影响
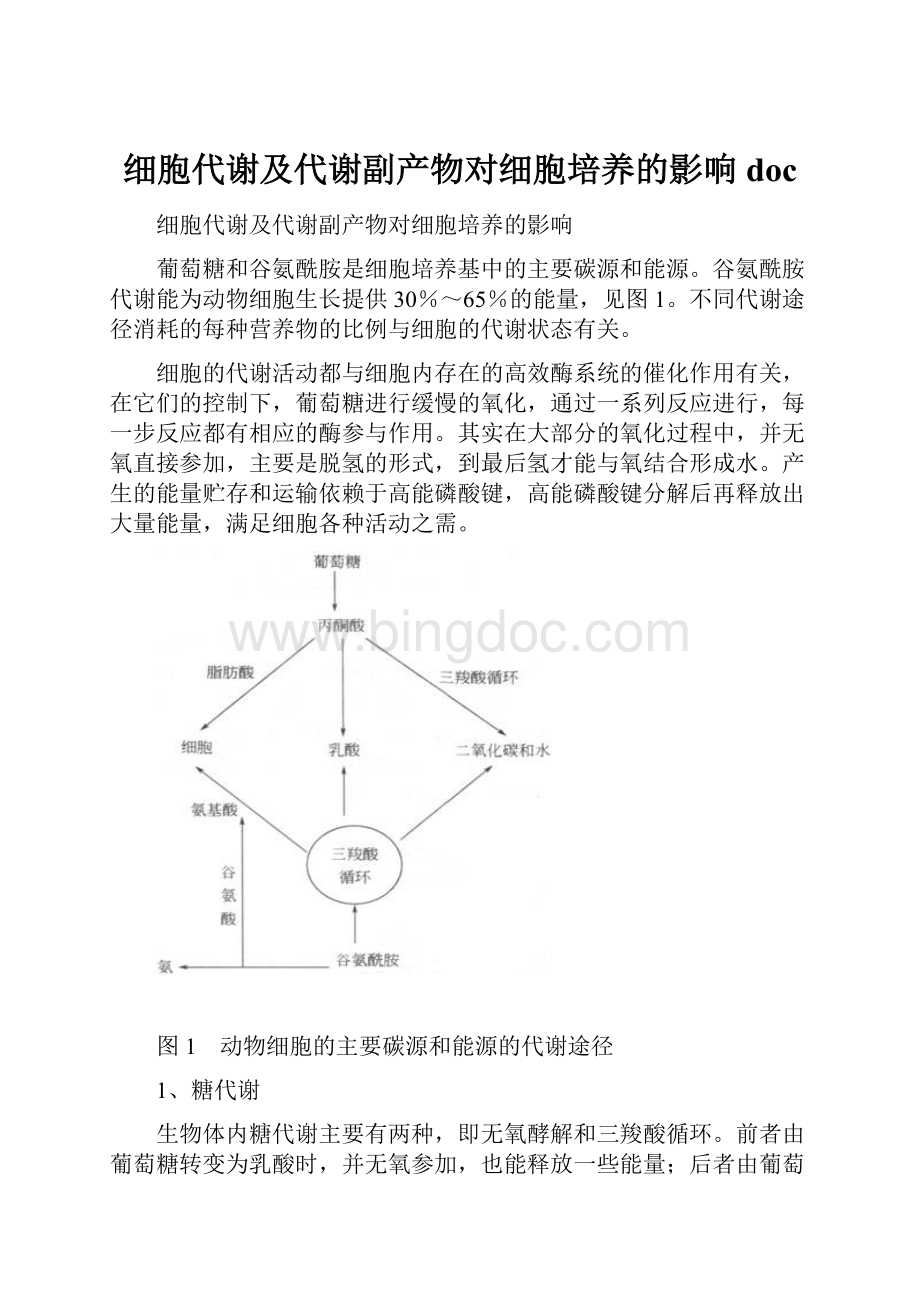
葡萄糖和谷氨酰胺是细胞培养基中的主要碳源和能源。
谷氨酰胺代谢能为动物细胞生长提供30%~65%的能量,见图1。
不同代谢途径消耗的每种营养物的比例与细胞的代谢状态有关。
细胞的代谢活动都与细胞内存在的高效酶系统的催化作用有关,在它们的控制下,葡萄糖进行缓慢的氧化,通过一系列反应进行,每一步反应都有相应的酶参与作用。
其实在大部分的氧化过程中,并无氧直接参加,主要是脱氢的形式,到最后氢才能与氧结合形成水。
产生的能量贮存和运输依赖于高能磷酸键,高能磷酸键分解后再释放出大量能量,满足细胞各种活动之需。
图1 动物细胞的主要碳源和能源的代谢途径
1、糖代谢
生物体内糖代谢主要有两种,即无氧酵解和三羧酸循环。
前者由葡萄糖转变为乳酸时,并无氧参加,也能释放一些能量;后者由葡萄糖转化为CO2和H2O时,需消耗大量的氧,但比前者产生的能量多。
1.1糖酵解途径
1分子的葡萄糖通过无氧酵解可净生成2个分子三磷酸腺苷(ATP),
这一过程全部在胞浆中完成。
在无氧情况下,葡萄糖分解生成乳酸的过程,叫做糖酵解。
糖酵解分三个阶段:
(1)第一阶段:
引发阶段。
由葡萄糖磷酸化、异构化、再磷酸化生成1,6-二磷酸果糖:
①葡萄糖磷酸化成为6-磷酸-葡萄糖,由己糖激酶催化。
为不可逆的磷酸化反应,酵解过程关键步骤之一,是葡萄糖进入任何代谢途径的起始反应,消耗1分子ATP。
②6-磷酸-葡萄糖转化为6-磷酸-果糖,磷酸己糖异构酶催化;
③6-磷酸-果糖磷酸化,转变为1,6-二磷酸-果糖;由6-磷酸果糖激酶催化,消耗1分子ATP,是第二个不可逆的磷酸化反应,酵解过程关键步骤之二,是葡萄糖氧化过程中最重要的调节点。
(2)第二阶段:
裂解阶段。
1,6-二磷酸-果糖折半分解成2分子磷酸丙糖(磷酸二羟丙酮和3-磷酸甘油醛),醛缩酶催化,二者可互变,最终1分子葡萄糖转变为2分子3-磷酸甘油醛。
(3)第三阶段:
通过氧化还原生成乳酸。
(能量的释放和保留)
①3-磷酸甘油醛的氧化和NAD+的还原,由3-磷酸甘油醛脱氢酶催化,生成1,3-二磷酸甘油酸,产生一个高能磷酸键,同时生成NADH用于第七步丙酮酸的还原。
②1,3-二磷酸甘油酸的氧化和ADP的磷酸化,生成3-磷酸甘油酸和ATP。
磷酸甘油酸激酶催化。
③3-磷酸甘油酸转变为2-磷酸甘油酸。
④2-磷酸甘油酸经烯醇化酶催化脱水,通过分子重排,生成具有一个高能磷酸键的磷酸烯醇式丙酮酸。
⑤磷酸烯醇式丙酮酸经丙酮酸激酶催化将高能磷酸键转移给ADP,生成烯醇式丙酮酸和ATP,为不可逆反应,酵解过程关键步骤之三。
⑥烯醇式丙酮酸与酮式丙酮酸的互变。
⑦丙酮酸还原生成乳酸。
反应中的NADH+H+来自于上述第1步反应中的3-磷酸甘油醛脱氢反应。
1.2TCA循环
在有氧的情况下,丙酮酸进入线粒体氧化脱羧生成的乙酰辅酶A,乙酰辅酶A与草酰乙酸缩合生成含有3个羧基的柠檬酸,再经过一系列反应重新变成草酰乙酸完成一轮循环(TCA循环),其中氧化反应脱下的氢经线粒体内膜上经呼吸链传递生成水,氧化磷酸化生成ATP;而脱羧反应生成的二氧化碳则通过血液运输到呼吸系统而被排出,是体内二氧化碳的主要来源。
在体外培养细胞中,葡萄糖经有氧氧化分解的比例取决于培养条件和培养细胞的特性。
三羧酸循环反应过程:
(1)乙酰辅酶A与草酰乙酸缩合生成柠檬酸
此反应由柠檬酸合酶(citratesynthase)催化,是三羧酸循环的关键酶,是重要的调节点。
由于高能硫酯键水解时释出较多自由能,ΔG'0=-32.2kJ/mol,此反应不可逆。
(2)柠檬酸经顺乌头酸生成异柠檬酸
此反应由顺乌头酸酶催化,柠檬酸脱水、加水生成异柠檬酸。
(3)异柠檬酸β-氧化、脱羧生成α-酮戊二酸
此反应在异柠檬酸脱氢酶作用下进行脱氢、脱羧,这是三羧酸循环中第一次氧化脱羧。
异柠檬酸脱氢酶(isocitratedehydrogenase)是三羧酸循环的限速酶,是最主要的调节点,辅酶是NAD+,脱氢生成的NADH+H+经线粒体内膜上经呼吸链传递生成水,氧化磷酸化生成3分子ATP。
异柠檬酸先脱氢生成草酰琥珀酸,再脱羧生成α-酮戊二酸。
ΔG'0=-20.9kJ/mol。
(4)α-酮戊二酸氧化、脱羧生成琥珀酰辅酶A
此反应在α-酮戊二酸脱氢酶复合体(α-ketoglutaratedehydrogenasecomplex)的催化下脱氢、脱羧生成琥珀酰辅酶A,这是三羧酸循环中第二次氧化脱羧。
α-酮戊二酸脱氢酶复合体是三羧酸循环的关键酶,是第三个调节点。
α-酮戊二酸脱氢酶复合体是多酶复合体,其组成及反应方式都与丙酮酸脱氢酶复合体相似。
它所含的三种酶是α-酮戊二酸脱氢酶(需TPP);硫辛酸琥珀酰基转移酶(需硫辛酸和辅酶A);二氢硫辛酸脱氢酶(需FAD、NAD+)。
脱氢生成NADH+H+,经线粒体内膜上经呼吸链传递生成水,氧化磷酸化生成3分子ATP。
由于反应中分子内部能量重排,产物琥珀酰辅酶A中含有一个高能硫酯键,此反应不可逆。
ΔG'0=-33.5kJ/mol。
(5)琥珀酰辅酶A转变为琥珀酸
此反应由琥珀酸硫激酶(琥珀酰辅酶A合成酶)催化,琥珀酰辅酶A中的高能硫酯键释放能量,可以转移给ADP(或GDP),形成ATP(或GTP)。
细胞中有两种同工酶,一种形成ATP,另一种形成GTP。
这是因为琥珀酸硫激酶由α、β亚基组成,α亚基上有磷酸化的组氨酸残基以及结合CoA的位点;β亚基上既可以结合ATP又可以结合GTP。
形成的GTP可在二磷酸核苷激酶催化下,将高能磷酸基团转移给ADP生成ATP。
这是三羧酸循环中唯一的一次底物水平磷酸化,生成1分子ATP。
(6)琥珀酸脱氢转变为延胡索酸
此反应由琥珀酸脱氢酶催化,辅酶是FAD,脱氢后生成FADH2,经线粒体内膜上经呼吸链传递生成水,氧化磷酸化生成2分子ATP。
(7)延胡索酸转变为苹果酸
此反应由延胡索酸酶催化,加水生成苹果酸。
(8)苹果酸脱氢生成草酰乙酸
此反应由苹果酸脱氢酶催化,辅酶是NAD+,脱氢后生成NADH+H+,经线粒体内膜上经呼吸链传递生成水,氧化磷酸化生成3分子ATP。
1.3葡萄糖糖代谢副产物对细胞培养的影响
(1)乳酸的来源:
乳酸是在培养过程中动物细胞代谢葡萄糖产生的主要代谢产物之一,对细胞的生长、代谢和产物的合成有着重要的影响。
乳酸主要来源于葡萄糖的代谢过程,谷氨酰胺及其他氨基酸的代谢也能产生少量乳酸(这部分乳酸约占乳酸总量的10%左右)。
在有氧的条件下,依细胞类型和培养条件的不同,葡萄糖转变成乳酸的量在5%~10%之间,最大可达70%。
有人证明,高压氧对胚胎组织或单层细胞培养都是有害的,而培养成体组织则需较多的氧。
CO2是葡萄糖代谢的最终产物,但它仍可被利用和丙酮酸一起形成草酸,重新进入三羧酸循环并起到增效作用。
CO2更重要的一个作用是它能调节细胞生存环境的pH值。
(2)乳酸对细胞生长代谢的影响
乳酸对细胞生长的不良影响主要是改变培养环境的pH和渗透压,间接的影响细胞的生长、代谢和产物的合成。
尽管不同的细胞系对乳酸具有不同的耐受能力,但只要维持pH值恒定,一般均能耐受较高浓度的乳酸。
在pH恒定的情况下,乳酸对细胞生长的影响可能来自两个方面,一是乳酸本身对细胞生理生化的影响;二是因乳酸的添加而导致了培养基渗透压的增加,继而影响细胞生长。
孙祥明等研究了乳酸对重组CHO细胞生长代谢及EPO表达的影响,发现:
①乳酸对细胞生长代谢和EPO表达量的影响的主要原因是渗透压的增加,乳酸分子本身对细胞生长代谢和EPO表达量没有明显作用。
②细胞的比生长速率随着乳酸浓度的增加而降低的主要原因是渗透压的作用而非乳酸分子本身,乳酸浓度从0增加到69mmol/L(对应渗透压300增加到420Osm/kg,见表1),细胞的比生长速率下降了33%。
③一般细胞适宜的渗透压范围是280~320Osm/kg,更高的渗透压通常会使细胞的生长速率减慢,但也有报道,高的渗透压可提高某些细胞株的产物合成速率。
乳酸浓度小于,对细胞生长的影响较小。
当乳酸浓度高于30mmol/L时,对细胞的影响较大,细胞存活率在培养过程中快速下降。
(3)乳酸对EPO表达量的影响
当乳酸浓度较大时,会使渗透压增大,而在高渗透压下细胞的比生长速率下降,从而造成EPO表达下降。
但尚不能排除渗透压对蛋白表达后的加工修饰、运输和分泌的影响,这有待于进一步的研究。
.
孙祥明等的乳酸和渗透压脉冲实验结果表明:
乳酸浓度从0增加到69mmol/L-1×10-3IU·cell-1·d-1下降到×10-3IU·cell-1·d-1。
如表4
(4)乳酸对EPO糖基化的影响
EPO的唾液酸含量随着,乳酸浓度的增加而增加(见表3)。
乳酸对寡糖链的种类没有明显影响。
乳酸对EPO糖基化的影响,可能是因为:
乳酸改变培养环境的pH,较低的pH更有利于EPO的糖基化。
2、谷氨酰胺代谢
葡萄糖和谷氨酰胺是细胞培养基中的主要碳源和能源。
谷氨酰胺是动物细胞培养中最重要的氨基酸,也是培养基中浓度最高的氨基酸,通常是其他氨基酸浓度的10倍左右。
谷氨酰胺是细胞生物合成中氮的主要来源,核酸中的嘌呤、嘧啶和氨基糖中的氮来自谷氨酰胺。
谷氨酰胺也可以作为碳源直接与细胞大分子物质的合成。
谷氨酰胺的存在可以促进其他氨基酸的运输和利用。
谷氨酰胺也是细胞培养中重要的能源物质。
谷氨酰胺代谢能为动物细胞生长提供30%~65%的能量。
不同代谢途径消耗的每种营养物的比例与细胞的代谢状态有关。
在细胞培养中,谷氨酰胺不稳定,容易降解。
细胞也会过量利用谷氨酰胺,导致氨的积累。
控制培养基中谷氨酰胺的浓度,可以有效较低氨的积累,改善细胞的生长环境。
在许多细胞培养过程中,均发现了细胞生长对谷氨酰胺有依赖性。
在杂交瘤细胞的培养中,谷氨酰胺的浓度为零时,细胞快速死亡。
重组CHO细胞能在无谷氨酰胺得培养基中生长,但生长速率减慢。
细胞生长是否依赖于谷氨酰胺的关键在于细胞是否含有内源的谷氨酰胺合成酶,即是否能将谷氨酸转化生成谷氨酰胺的能力,谷氨酰胺的运输速率及谷氨酰胺的生成速率限制是导致细胞在无谷氨酰胺培养基中生长速率减慢的主要原因。
谷氨酰胺作为能源物质,通过转氨反应或者脱氨反应生成α-酮戊二酸,进入三羧酸循环。
反应的第一步是在磷依赖的谷氨酰胺酶(PAG)的作用下,脱去酰胺基,生成谷氨酸。
反应的第二步是谷氨酸到α-酮戊二酸的转化,是谷氨酰胺代谢的关键步骤,普遍认为存在反应方式的选择问题,谷氨酸可以在谷氨酸脱氢酶的作用下生成α-酮戊二酸(每摩尔的谷氨酸可以生成1mol的氨),也可以在谷草转氨酶或丙转氨酶的作用下,将氨基转给草酰乙酸或丙酮酸,分别生成天冬氨酸或丙氨酸。
代谢途径的选择取决于细胞株的本性和培养环境。
CHO细胞主要以转氨途径利用谷氨酸。
谷氨酰胺的胞内代谢途径如下图:
(1)氨的来源
氨主要是由谷氨酰胺等氨基酸脱氨产生的。
细胞培养过程中,谷氨酰胺的消耗比其它氨基酸的总和还多,80%~90%的氨都是由谷氨酰胺代谢产生的。
(2)氨对细胞生长代谢的影响
氨也是细胞培养过程中的主要代谢副产物,与乳酸相比,较低浓度的氨就会对细胞生长产生抑制。
氨的积累使细胞内的UDP氨基己糖增加,影响细胞的生长及蛋白质的糖基化过程。
氨抑制谷氨酰胺代谢途径,使天冬氨酸和谷氨酸的消耗增加,影响细胞的氨基代谢。
同时氨浓度的提高,改变了细胞内局部微环境的pH,影响细胞的正常生理功能。
不同的细胞株对氨有不同的耐受能力。
氨对细胞的生长、代谢和重组蛋白的表达都有影响。
氨不仅对细胞的生长具有较强的抑制作用,而且对细胞的生存也具有毒性。
当氨的浓度高于3mmol/L时,对细胞的生长具有明显的抑制作用。
氨对细胞生长的影响,主要是影响了细胞的代谢。
乳酸对葡萄糖的得率系数随着氨浓度的增加而增加,这表明,氨浓度的提高导致葡萄糖代谢更倾向于乳酸的生成,在消耗掉的葡萄糖中,经厌氧代谢消耗的葡萄糖的比例随着氨浓度的提高而提高。
但不同的细胞株,氨对葡萄糖代谢有着不同的影响机理。
(3)氨对EPO表达量的影响
目前研究氨对EPO表达量的影响,都是在培养基中加入不同浓度的氨。
研究表明:
EPO表达量随着氨浓度的增加(0.21~5.66mmol/L)而下降,主要是因为氨对细胞生长具有抑制作用,导致最终细胞密度随着氨浓度的增加而下降。
但张兴元等研究了氨浓度对产物比生成速率的影响,结果表明:
EPO的比生成速率首先随着氨浓度的增加(1.85~3.22mmol/L)而增加,进而又随着氨浓度的进一步增加(3.22~7.22mmol/L)而下降。
3、代谢调控培养
乳酸和氨是在培养过程中动物细胞产生的主要代谢产物,对细胞的正常生理功能在抑制甚至毒害作用。
在分批培养和补料分批培养中的这一问题尤为突出。
尽管灌注培养可以通过提高灌注速度来去除抑制产物。
但是,一方面由于灌注培养细胞浓度很高,提高灌注速率,营养成分的供给十分充分,氨和乳酸的产生速率也增加了。
另一方面,过高的灌注速率提高了细胞的比生长速率,降低产物的比产率,加上细胞对营养的利用并不彻底,培养液中会残留大量的蛋白,造成提取纯化的不便和培养基的浪费。
所以,在培养中调控动物细胞的代谢途径一直较受重视。
通过代谢调控,可以减少副产物的产生,降低细胞的死亡速率,还可以控制灌注速率和培养液成分,控制细胞的状态和比生长速率,以提高目标蛋白的产率。
大量实验表明,细胞的比生长速率降低时,产物的比生长速率提高,有人控制细胞的比生长速率为最大比生长速率的60%抗体生长速率增加了97%。
具体有以下方法。
3.1 控制葡萄糖浓度法
乳酸浓度升高,会导致比生长速率降低,比死亡速率升高。
乳酸的降低可更换葡萄糖为己糖如果糖或半乳糖,还可限制葡萄糖减少乳酸的生成,使初始葡萄糖浓度较低,在培养过程中再添加。
在控制葡萄糖浓度法培养中,生长期可以使葡萄糖浓度稍高,以促进细胞生长;在产物合成期降低葡萄糖的浓度,降低乳酸的产生速率,避免乳酸的积累,减少毒害,降低死亡速率,维护持活细胞数在较高水平。
同时还可以降低比生长速率,增加目标蛋白的产生速率。
灌注葡萄糖的同时,要间歇或连续地加入其他组分,以避免营养缺乏,其中谷氨酰胺要保持在较低水平,因为细胞的生长不依赖糖酵解,即使没有葡萄糖,细胞仍可以通过降解谷氨酰胺获得能量。
假如谷氨酰胺的浓度过高,细胞就会偏向谷氨酰胺酵解,从而削弱这种方法的效果。
由于葡萄糖的价格相对低廉,这种方法很有前途。
3.2 控制谷氨酰胺法
上面提到,没有葡萄糖,细胞可以利用谷氨酰胺作能量物质。
因此,控制谷氨酰胺比控制葡萄糖要容易些,应用这一方法的报道也较多。
控制谷氨酰胺浓度的目的,主要是减少氨的产生。
氨对细胞的毒性比乳酸大得多,表现为降低比生长速率,增加死亡速率。
有人详细地研究了动物细胞的代谢过程,采用底物限制补料分批工艺对动物细胞进行代谢控制。
他采用这一方法,使氨的浓度降低了一半。
控制谷氨酰胺法与控制葡萄糖法一样,要维持葡萄糖在较低水平。
3.3 控制葡萄糖和谷氨酰胺法
在细胞中,葡萄糖代谢和谷氨酰胺代谢密切相关。
葡萄糖消耗上升,则谷氨酰胺消耗下降,反之亦然。
在相当大的一个范围内,葡萄糖和谷氨酰胺的消耗速率与其浓度成正比。
控制葡萄糖和谷氨酰胺法可降低乳酸和氨的产生,还能有效控制比生长速率。
在细胞生长期,提供充分的营养,供细胞的需要;在产物合成期,降低葡萄糖和谷氨酰胺的浓度或流量,降低比生长速率,增加目标蛋白的产率。
3.4 代谢产物的去除
通常使用透析膜,超滤腊或吸附剂选择性去除乳酸、氨或铵离子。
有人建议加化学试剂比如钾盐来消除氨的影响,也有人建议可使用有谷氨酰胺合成酶的细胞。
灌注培养发展到现在,还有许多急待解决的问题。
其最大的缺点是培养基的利用不充分,造成一定的浪费。
随着细胞培养技术和产品分离技术的进一步发展,建立细胞培养与产物分离的耦合系统,能充分利用培养液,降低生产成本,一直是人们追求的目标。
小结:
1、乳酸对细胞生长代谢和EPO表达量的影响的主要原因是造成渗透压的增加,高渗透压下细胞的比生长速率下降,从而造成EPO表达下降。
因此,发酵过程在pH恒定时,细胞可以耐受较高浓度的乳酸,乳酸浓度小于,对细胞生长的影响较小。
当乳酸浓度高于30mmol/L时,对细胞的影响较大,细胞存活率在培养过程中快速下降。
2、与乳酸相比,较低浓度的氨就会对细胞生长产生抑制。
当氨的浓度高于3mmol/L时,就会对细胞的生长产生明显的抑制作用。
因此,发酵过程通过控制培养基中谷氨酰胺的浓度,可以有效较低氨的积累,改善细胞的生长环境。
3、在CHO细胞中,谷氨酰胺浓度对谷氨酰胺的消耗、氨和丙氨酸的生成均有较大的影响。
在谷氨酰胺限制流加培养过程中可以控制谷氨酰胺在较低浓度,改变了谷氨酰胺的代谢流量分布,可显著提高谷氨酰胺及其能量代谢的利用率,减少副产物氨和丙氨酸的生成和积累,从而有效解决营养物耗竭和代谢副产物积累的矛盾,提高动物细胞大规模培养过程的经济性。


- 配套讲稿:
如PPT文件的首页显示word图标,表示该PPT已包含配套word讲稿。双击word图标可打开word文档。
- 特殊限制:
部分文档作品中含有的国旗、国徽等图片,仅作为作品整体效果示例展示,禁止商用。设计者仅对作品中独创性部分享有著作权。
- 关 键 词:
- 细胞 代谢 副产物 细胞培养 影响 doc
 冰点文库所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
冰点文库所有资源均是用户自行上传分享,仅供网友学习交流,未经上传用户书面授权,请勿作他用。
 《保险学概论》期末复习指导书蓝本答案要点.docx
《保险学概论》期末复习指导书蓝本答案要点.docx
-
《读李白诗歌 品太白遗韵》.docx
-
《海底两万里》读书笔记800字.docx
-
《经济法基础》考试真题及答案921.docx
-
《石灰吟》教学设计部编版1.docx
-
《数字化测图》课程标准.docx
-
#网上购物系统需求分析.docx
-
《大棚蔬菜》教案doc.docx
-
《各具特色的民居》综合资料.docx
-
#加强中小型企业人力资源管理的对策思考.docx
-
《计算机文化基础wordl》课程单元教学设计.docx
-
《不会叫的狗》教案 人教部编版1.docx
-
《菜根谭》注译四闲适篇.docx
-
《反不正当竞争法》下互联网平台封禁行为考辨以消费者用户合法权益保护为中心.docx
-
《福尔摩斯探案集》读后感15篇.docx
-
《荷叶圆圆》公开课教案优秀教学设计26.docx
-
《操作系统》二学期A卷及答案.docx
-
《科技出行研究报告》智能网联与新能源将变革未来汽车出行.docx
-
《红对勾》古代诗歌选择题答案补充.docx
-
《傅雷家书》名著阅读笔记.docx
-
《课堂密码》读后感及心得精选多篇.docx
-
《时间旅行者的妻子》中英文剧本.docx
-
《化工原理》第六章蒸发.docx
-
《你若安好便是晴天林徽因传》序.docx
-
《蓝海战略》概要11页.docx
-
《体育科学》论文格式.docx
-
《劝学师说赤壁赋登泰山记》文言现象翻译简答题.docx
-
《微观经济学》期末试题.docx
-
《微机测控系统原理与设计》习题解答.docx
-
《扫雪心得体会》.docx
-
《质量处理学》学习规划.docx
-
《物质的密度》教案.docx
-
专题13 状语从句解析版.docx
-
渝建竣资料全套表格.docx
-
语文命题培训材料.docx
-
育婴员中级操作技能复习提纲0.docx
-
园林树木学实验树种.docx
-
原始凭证的审核研究.docx
-
云南民族大学论文撰写规范.docx
-
再见了我亲爱的母校作文.docx
-
在学习贯彻中组部干部选拔任用工作视频会议精神会议上的讲话.docx
-
怎样选择适合你的主板最全主板选购指南.docx
-
张掖丹霞地貌解析.docx
-
招聘面试案例.docx
-
浙江省宁波市届高三下学期适应性模拟考试语文试题.docx
-
真情义卖策划书1.docx
-
整理建筑工程答案与题目89.docx
-
证券从业资格考试证券投资顾问考前冲刺试题1doc.docx
-
执业医师技能考试评分标准.docx
-
职业认知教学教案.docx
-
制冷实习报告4篇.docx
